Worldwide Web: High Venom Potency and Ability to Optimize Venom Usage Make the Globally Invasive Noble False Widow Spider Steatoda nobilis (Thorell, 1875) (Theridiidae) Highly Competitive against Native European Spiders Sharing the Same Habitats

 , ,
, ,
Abstract
:1. Introduction
2. Results
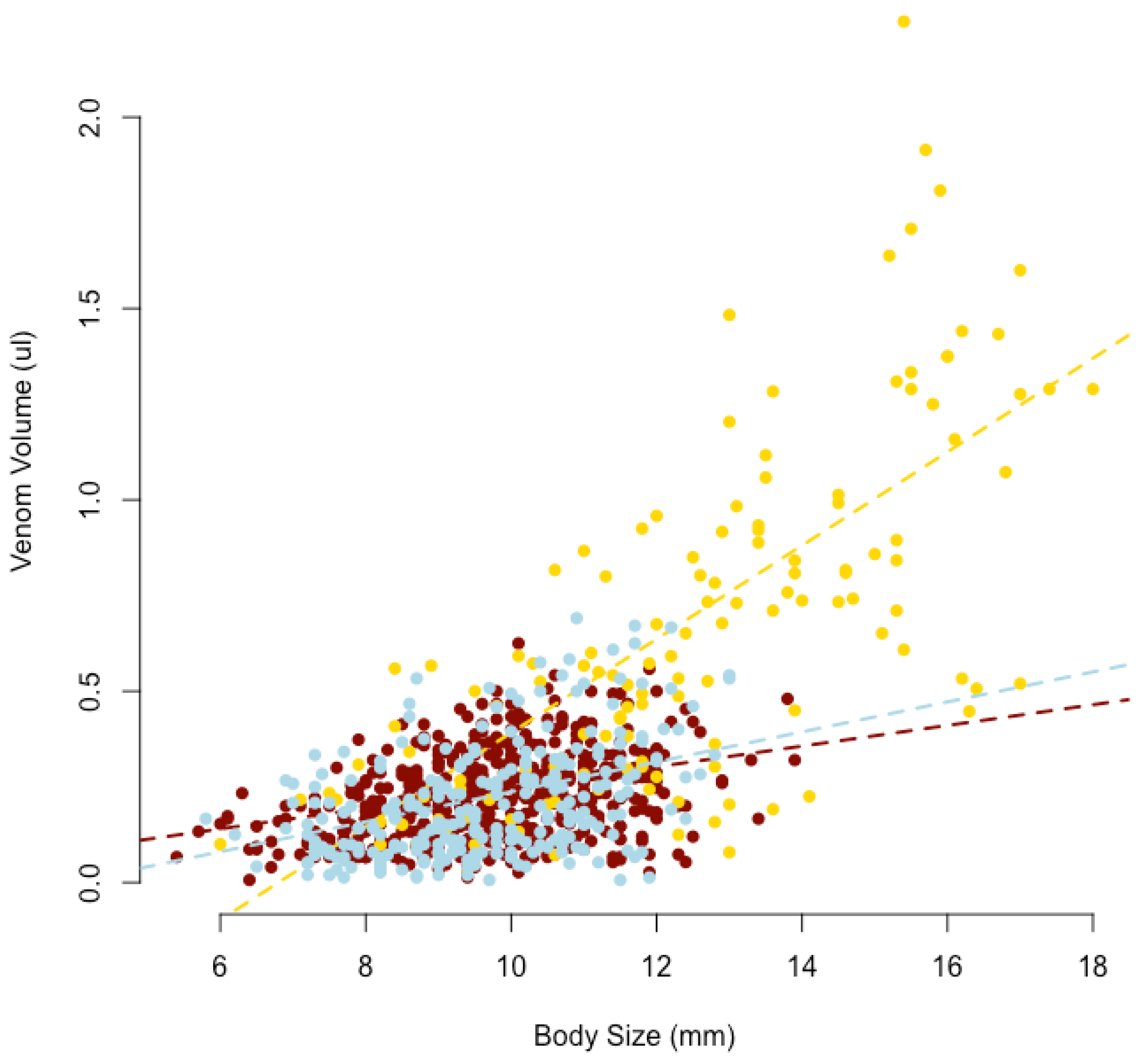
2.1. Spider Size and Venom Yield
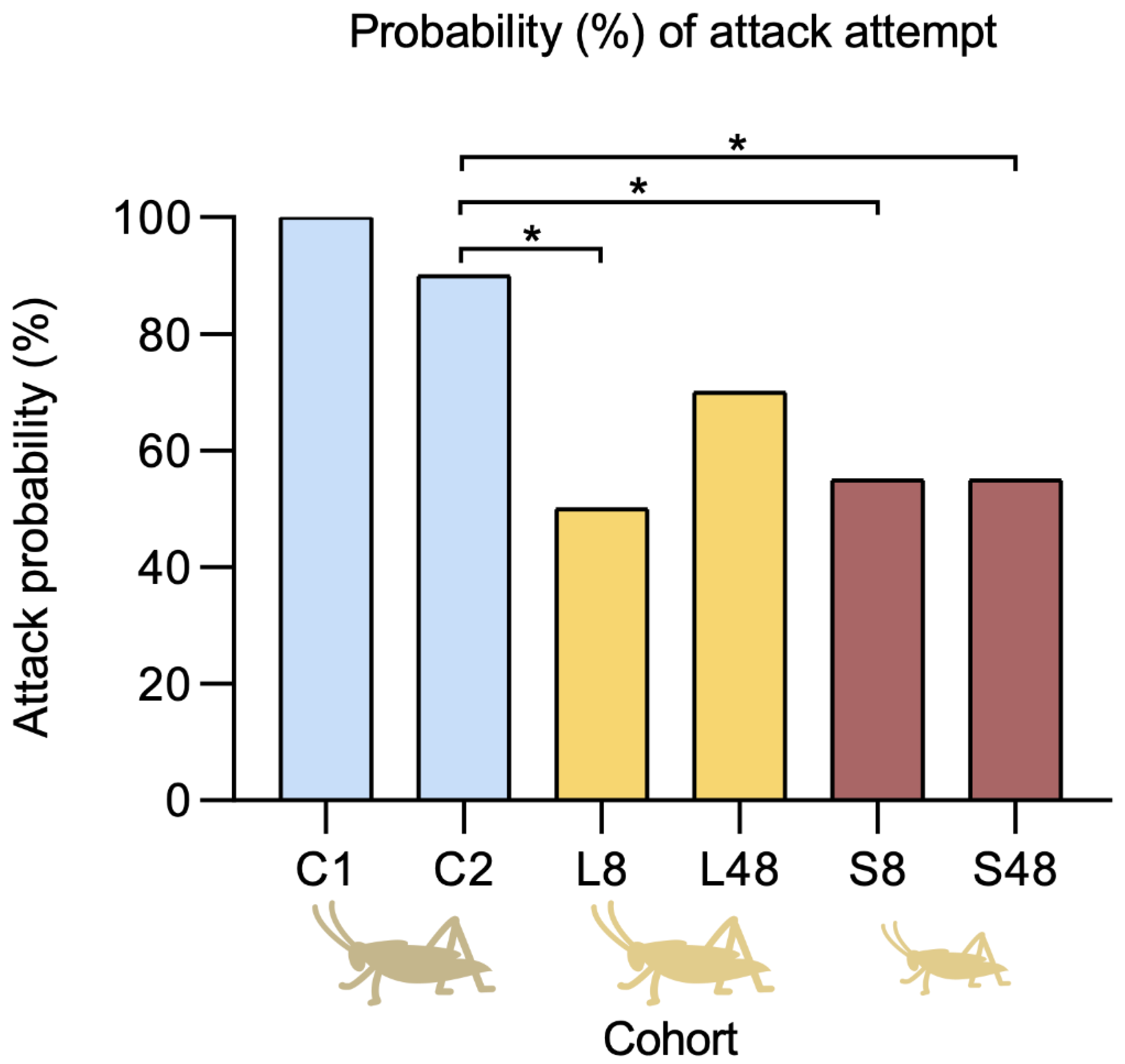
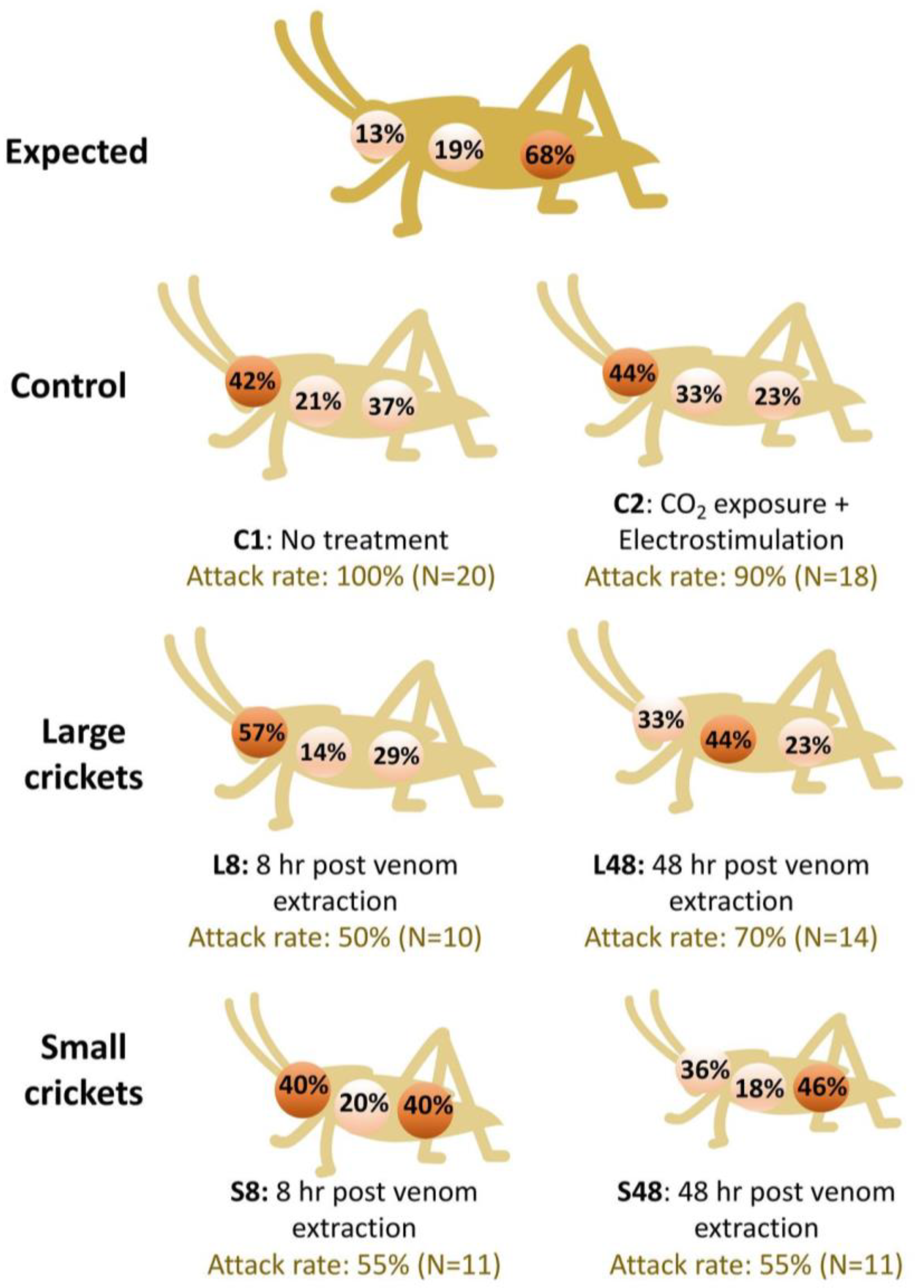
2.2. Predatory Behaviour Influenced by Venom Availability
2.3. S. nobilis, A. similis and E. atrica Venom Toxicity
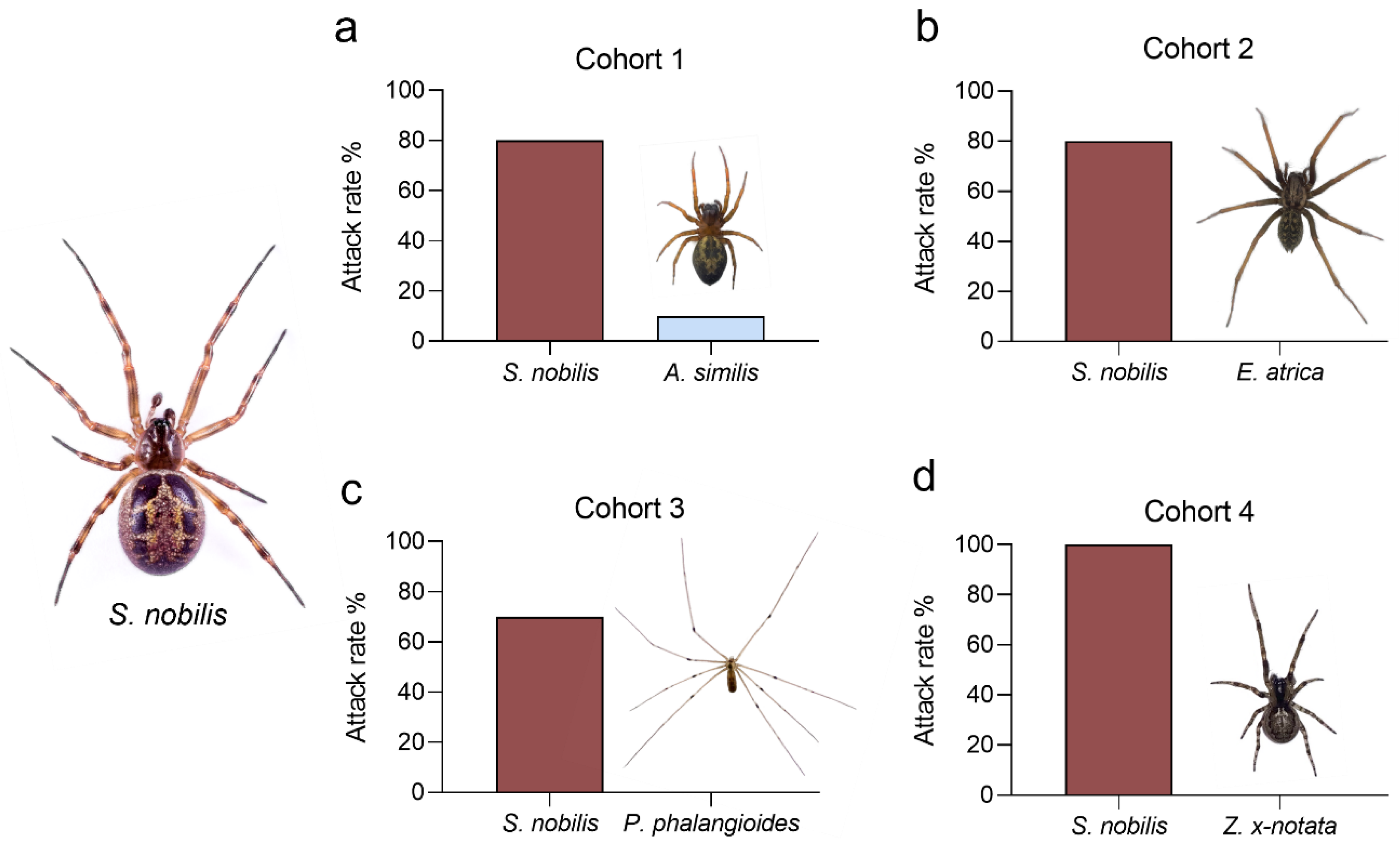
2.4. Predation Observations
3. Discussion
3.1. Predatory Behaviour Influenced by Venom Availability
3.2. Venom Yield and Potency
3.3. Competitiveness/Intraguild Predation
4. Materials and Methods
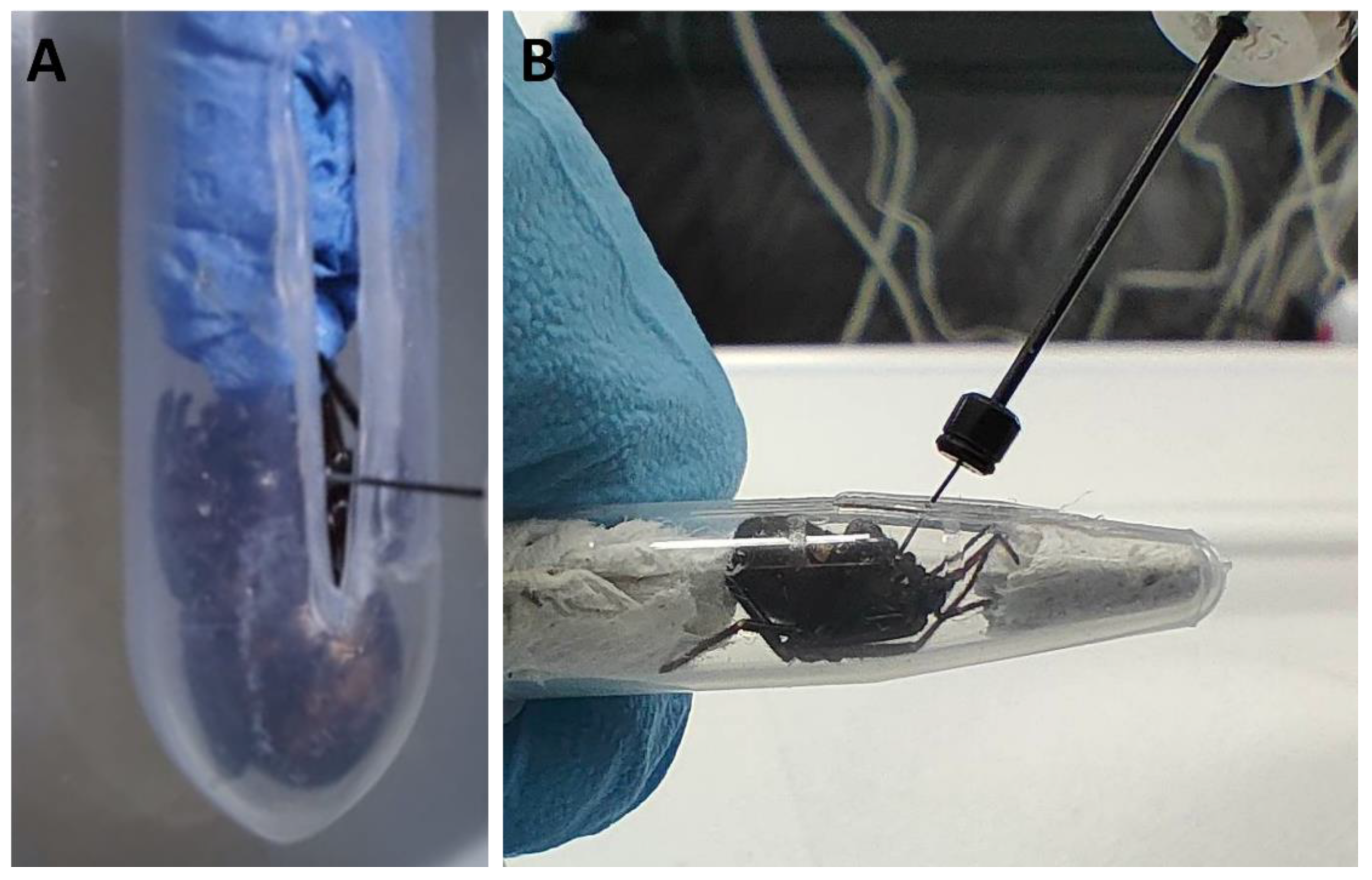
4.1. Spider Collection, Housing, Measurements, and Venom Extraction
4.2. Venom Toxicity Assays
4.3. Influence of Venom Availability on Behaviour
4.4. Predation Observations
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Spider Catalogue. World Spider Catalogue Version 23.5. Natural History Museum Bern. 2022. Available online: http://wsc.nmbe.ch (accessed on 4 August 2022).
- Nyffeler, M.; Birkhofer, K. An estimated 400–800 million tons of prey are annually killed by the global spider community. Sci. Nat. 2017, 104, 30. [Google Scholar] [CrossRef] [PubMed]
- Badenhausser, I.; Gross, N.; Mornet, V.; Roncoroni, M.; Saintilan, A.; Rusch, A. Increasing amount and quality of green infrastructures at different scales promotes biological control in agricultural landscapes. Agric. Ecosyst. Environ. 2019, 290, 106735. [Google Scholar] [CrossRef]
- Barin, A.; Arabkhazaeli, F.; Rahbari, S.; Madani, S.A. The housefly, Musca domestica, as a possible mechanical vector of Newcastle disease virus in the laboratory and field. Med. Vet. Entomol. 2010, 24, 88–90. [Google Scholar] [CrossRef] [PubMed]
- Ndava, J.; Llera, S.D.; Manyanga, P. The future of mosquito control: The role of spiders as biological control agents: A review. Int. J. Mosq. Res. 2018, 5, 6–11. [Google Scholar]
- Wang, Y.-C.; Chang, Y.-C.; Chuang, H.-L.; Chiu, C.-C.; Yeh, K.-S.; Chang, C.-C.; Hsuan, S.-L.; Lin, W.-H.; Chen, T.-H. Transmission of Salmonella between Swine Farms by the Housefly (Musca domestica). J. Food Prot. 2011, 74, 1012–1016. [Google Scholar] [CrossRef]
- Hann, S. Evidence for the displacement of an endemic New Zealand spider, Latrodectus katipo Powell by the South African species Steatoda capensis Hann (Araneae: Theridiidae). N. Z. J. Zool. 1990, 17, 295–307. [Google Scholar] [CrossRef]
- Nyffeler, M.; Dondale, C.D.; Redner, J.H. Evidence for displacement of a North American spider, Steatoda borealis (Hentz), by the European species S. bipunctata (Linnaeus) (Araneae: Theridiidae). Can. J. Zool. 1986, 64, 867–874. [Google Scholar] [CrossRef]
- Bednarski, J.; Ginsberg, H.; Jakob, E.M. Competitive interactions between a native spider (Frontinella communis, Araneae: Linyphiidae) and an invasive spider (Linyphia triangularis, Araneae: Linyphiidae). Biol. Invasions 2010, 12, 905–912. [Google Scholar] [CrossRef]
- Rozwałka, R.; Rutkowski, T.; Bielak-Bielecki, P. New data on introduced and rare synanthropic spider species (Arachnida: Araneae) in Poland (II). Ann. Univ. Mariae Curie-Sklodowska Sect. C–Biol. 2017, 71, 59. [Google Scholar] [CrossRef]
- Suvák, M. Invasive spider Uloborus plumipes Lucas, 1846 (Araneae: Uloboridae), new to Slovakia. Folia Faun. Slovaca 2013, 18, 39–45. [Google Scholar]
- Taucare-Ríos, A.; Mardones, D.; Zúñiga-Reinoso, A. Steatoda nobilis (Araneae: Theridiidae) in South America: A new alien species for Chile. Can. Entomol. 2016, 148, 479–481. [Google Scholar] [CrossRef]
- Kumschick, S.; Fronzek, S.; Entling, M.H.; Nentwig, W. Rapid spread of the wasp spider Argiope bruennichi across Europe: A consequence of climate change? Clim. Change 2011, 109, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Krehenwinkel, H.; Rödder, D.; Năpăruş-Aljančič, M.; Kuntner, M. Rapid genetic and ecological differentiation during the northern range expansion of the venomous yellow sac spider Cheiracanthium punctorium in Europe. Evol. Appl. 2016, 9, 1229–1240. [Google Scholar] [CrossRef]
- Vink, C.J.; Derraik, J.; Phillips, C.B.; Sirvid, P.J. The invasive Australian redback spider, Latrodectus hasseltii Thorell 1870 (Araneae: Theridiidae): Current and potential distributions, and likely impacts. Biol. Invasions 2011, 13, 1003–1019. [Google Scholar] [CrossRef]
- Harvey, P. Zoropsis spinimana (Dufour, 1820) established indoors in Britain. Newsl. Br. Arachnol. Soc. 2012, 112, 20–21. [Google Scholar]
- Nadolny, A.A. The first record of Zoropsis spinimana (Aranei, Zoropsidae) in the Crimea. Zool. Ecol. 2016, 26, 127–128. [Google Scholar] [CrossRef]
- Purgat, P.; Ondrejková, N.; Krumpálová, Z.; Gajdoš, P.; Hurajtová, N. Tegenaria hasperi Chyzer, 1897 and Zoropsis spinimana (Dufour, 1820), newly recorded synanthropic spiders from Slovakia (Araneae, Agelenidae, Zoropsidae). Check List 2021, 17, 775–782. [Google Scholar] [CrossRef]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Mooney, H.A.; Hobbs, R.J. Global change and invasive species: Where do we go from here. In Invasive Species in a Changing World; Island Press: Washington, DC, USA, 2000; pp. 425–434. [Google Scholar]
- Kobelt, M.; Nentwig, W. Alien spider introductions to Europe supported by global trade. Divers. Distrib. 2007, 14, 273–280. [Google Scholar] [CrossRef]
- Nentwig, W.; Pantini, P.; Vetter, R.S. Distribution and medical aspects of Loxosceles rufescens, one of the most invasive spiders of the world (Araneae: Sicariidae). Toxicon 2017, 132, 19–28. [Google Scholar] [CrossRef]
- Dugon, M.M.; Dunbar, J.P.; Afoullouss, S.; Schulte, J.; McEvoy, A.; English, M.J.; Hogan, R.; Ennis, C.; Sulpice, R. Occurrence, reproductive rate and identification of the non-native Noble false widow spider Steatoda nobilis (Thorell, 1875) in Ireland. Boil. Environ. Proc. R. Ir. Acad. 2017, 117B, 77. [Google Scholar] [CrossRef]
- Dunbar, J.; Ennis, C.; Gandola, R.; Dugon, M. Biting off more than one can chew: First record of the non-native Noble false widow spider Steatoda nobilis (Thorell, 1875) feeding on the native Viviparous lizard Zootoca vivipara. Biol. Environ. Proc. R. Ir. Acad. 2018, 118, 45–48. [Google Scholar] [CrossRef]
- Dunbar, J.P.; Vitkauskaite, A.; Lawton, C.; Waddams, B.; Dugon, M.M. Webslinger versus dark knight: First record of a false widow spider Steatoda nobilis preying on a pipistrelle bat in Britain. Ecosphere 2022, 13, e3959. [Google Scholar] [CrossRef]
- Faúndez, E.; Johnson, E.; Angelone, E.V. A case of predation by the noble false widow Steatoda nobilis (Thorell, 1875) (Araneae: Theridiidae) on the small milkweed bug Lygaeus kalmii kalmii Stal, 1874 (Heteroptera: Lygaeidae). Revista Ibérica de Aracnología 2020, 37, 275–277. [Google Scholar]
- Locket, G.H. Some notes on the life history of Steatoda nobilis (Thorell). Newsl. Br. Arachnol. Soc. 1979, 25, 8–10. [Google Scholar]
- Thorell, T. Diagnoses aranearum Europaearum aliquot novarum. Tijdschrift voor Entomologie 1875, 18, 81–108. [Google Scholar]
- Snazell, R.; Jones, D. The theridiid spider Steatoda nobilis (Thorell, 1875) in Britain. Bull. Br. Arachnol. Soc. 1993, 9, 164–167. [Google Scholar]
- Bauer, T.; Feldmeier, S.; Krehenwinkel, H.; Wieczorrek, C.; Reiser, N.; Breitling, R. Steatoda nobilis, a false widow on the rise: A synthesis of past and current distribution trends. NeoBiota 2019, 42, 19. [Google Scholar] [CrossRef]
- Hambler, C. The Noble false widow spider Steatoda nobilis is an emerging public health and ecological threat. OSF Prepr. 2019, axbd4. [Google Scholar] [CrossRef]
- Dunbar, J.P.; Schulte, J.; Lyons, K.; Fort, A.; Dugon, M.M. New Irish record for Steatoda triangulosa (Walckenaer, 1802), and new county records for Steatoda nobilis (Thorell, 1875), Steatoda bipunctata (Linnaeus, 1758) and Steatoda grossa (CL Koch, 1838). Ir. Nat. J. 2018, 36, 39–43. [Google Scholar]
- Faúndez, E.I.; Téllez, F. Primer registro de una mordedura de Steatoda nobilis (Thorell, 1875) (Arachnida: Araneae: Theridiidae) en Chile. Arquivos Entomolóxicos 2016, 15, 237–240. [Google Scholar]
- Vetter, R.S.; Adams, R.J.; Berrian, J.E.; Vincent, L. The European spider Steatoda nobilis (Thorell, 1875) (Araneae: Theridiidae) becoming widespread in California. Pan-Pac. Entomol. 2015, 91, 98–101. [Google Scholar] [CrossRef]
- Vetter, R.S.; Rust, M.K. A large European combfoot spider, Steatoda nobilis (Thorell 1875) (Araneae: Theridiidae), newly established in Ventura County, California. Pan-Pac. Entomol. 2012, 88, 92–98. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Omran, M.A.A.; Abdel-Nabi, I.M.; Ueda, H.; McVean, A. Intraspecific variation in the Egyptian scorpion Scorpio maurus palmatus venom collected from different biotopes. Toxicon 2009, 53, 349–359. [Google Scholar] [CrossRef]
- Zamani, A.; Mirshamsi, O.; Jannesar, B.; Marusik, Y.M.; Esyunin, S.L. New data on spider fauna of Iran (Arachnida: Araneae), Part II. Zool. Ecol. 2017, 25, 339–346. [Google Scholar] [CrossRef]
- Nolan, M. Three Spiders (Araneae) New to Ireland: Bolyphantes alticeps, Oonops domesticus and Steatoda nobilis. Ir. Nat. J. 1999, 26, 200–202. [Google Scholar]
- Dunbar, J.P.; Khan, N.A.; Abberton, C.L.; Brosnan, P.; Murphy, J.; Afoullouss, S.; O’Flaherty, V.; Dugon, M.M.; Boyd, A. Synanthropic spiders, including the global invasive noble false widow Steatoda nobilis, are reservoirs for medically important and antibiotic resistant bacteria. Sci. Rep. 2020, 10, 20916. [Google Scholar] [CrossRef]
- Faúndez, E.I.; Carvajal, M.A.; Aravena-Correa, N.P. On a bite by Steatoda nobilis (Thorell, 1875) (Araneae: Theridiidae) on a human being, with comments on its handling during the 2020 SARS-COV-2 pandemic. Rev. Iber. Aracnol. 2020, 51, 178–180. [Google Scholar]
- Warrell, D.; Shaheen, J.; Hillyard, P.; Jones, D. Neurotoxic envenoming by an immigrant spider (Steatoda nobilis) in southern England. Toxicon 1991, 29, 1263–1265. [Google Scholar] [CrossRef]
- Kulczycki, A.; Simeon, E.; Legittimo, C.M.; Di Pompeo, P. New Records of Steatoda nobilis (Thorell, 1875) (Araneae, Theridiidae), an Introduced Species on the Italian Mainland and in Sardinia. Arachnology 2012, 15, 269–272. [Google Scholar] [CrossRef]
- Dunbar, J.P.; Vitkauskaite, A.; O’Keeffe, D.T.; Fort, A.; Sulpice, R.; Dugon, M.M. Bites by the noble false widow spider Steatoda nobilis can induce Latrodectus-like symptoms and vector-borne bacterial infections with implications for public health: A case series. Clin. Toxicol. 2021, 60, 59–70. [Google Scholar] [CrossRef]
- Dunbar, J.P.; Fort, A.; Redureau, D.; Sulpice, R.; Dugon, M.M.; Quinton, L. Venomics Approach Reveals a High Proportion of Lactrodectus-Like Toxins in the Venom of the Noble False Widow Spider Steatoda nobilis. Toxins 2020, 12, 402. [Google Scholar] [CrossRef]
- Vitkauskaite, A.; Dunbar, J.P.; Lawton, C.; Dalagiorgos, P.; Allen, M.M.; Dugon, M.M. Vertebrate prey capture by Latrodectus mactans (Walckenaer, 1805) and Steatoda triangulosa (Walckenaer, 1802) (Araneae, Theridiidae) provide further insights into the immobilization and hoisting mechanisms of large prey. Food Webs 2021, 29, e00210. [Google Scholar] [CrossRef]
- Dunbar, J.P.; Sulpice, R.; Dugon, M.M. The kiss of (cell) death: Can venom-induced immune response contribute to dermal necrosis following arthropod envenomations? Clin. Toxicol. 2019, 57, 677–685. [Google Scholar] [CrossRef]
- Nelsen, D.R.; Kelln, W.; Hayes, W.K. Poke but don’t pinch: Risk assessment and venom metering in the western black widow spider, Latrodectus hesperus. Anim. Behav. 2014, 89, 107–114. [Google Scholar] [CrossRef]
- Wigger, E.; Kuhn-Nentwig, L.; Nentwig, W. The venom optimisation hypothesis: A spider injects large venom quantities only into difficult prey types. Toxicon 2002, 40, 749–752. [Google Scholar] [CrossRef]
- Hayes, W. The snake venom-metering controversy: Levels of analysis, assumptions, and evidence. In The Biology of Rattlesnakes; Loma Linda University Press: Loma Linda, CA, USA, 2008; pp. 191–220. [Google Scholar]
- Hayes, W.K. Venom metering by juvenile prairie rattlesnakes, Crotalus v. viridis: Effects of prey size and experience. Anim. Behav. 1995, 50, 33–40. [Google Scholar] [CrossRef]
- Hayes, W.K.; Herbert, S.S.; Rehling, G.; Gennaro, J. Factors that influence venom expenditure in viperids and other snake species during predatory and defensive contexts. In Biology of the Vipers; Eagle Mountain Publishing: Eagle Mountain, UT, USA, 2002; pp. 207–233. [Google Scholar]
- Young, B.A.; Lee, C.E.; Daley, K.M. Do snakes meter venom? BioScience 2002, 52, 1121–1126. [Google Scholar] [CrossRef]
- Dugon, M.M.; Arthur, W. Prey orientation and the role of venom availability in the predatory behaviour of the centipede Scolopendra subspinipes mutilans (Arthropoda: Chilopoda). J. Insect Physiol. 2012, 58, 874–880. [Google Scholar] [CrossRef]
- Edmunds, M.C.; Sibly, R.M. Optimal sting use in the feeding behavior of the scorpion Hadrurus spadix. J. Arachnol. 2010, 38, 123–125. [Google Scholar] [CrossRef]
- Lira, A.F.; Santos, A.B.; Silva, N.A.; Martins, R.D. Threat level influences the use of venom in a scorpion species, Tityus stigmurus (Scorpiones, Buthidae). Acta Ethol. 2017, 20, 291–295. [Google Scholar] [CrossRef]
- Nisani, Z.; Hayes, W.K. Defensive stinging by Parabuthus transvaalicus scorpions: Risk assessment and venom metering. Anim. Behav. 2011, 81, 627–633. [Google Scholar] [CrossRef]
- Cooper, A.M.; Nelsen, D.R.; Hayes, W.K. The Strategic Use of Venom by Spiders. In Toxinology: Evolution of Venomous Animal and Their Toxins; Springer: Dordrecht, The Netherlands, 2015; pp. 1–18. [Google Scholar]
- Hostettler, S.; Nentwig, W. Olfactory information saves venom during prey-capture of the hunting spider Cupiennius salei (Araneae: Ctenidae). Funct. Ecol. 2006, 20, 369–375. [Google Scholar] [CrossRef]
- Wullschleger, B.; Nentwig, W. Influence of venom availability on a spider’s prey-choice behaviour. Funct. Ecol. 2002, 16, 802–807. [Google Scholar] [CrossRef]
- Boevé, J.-L.; Kuhn-Nentwig, L.; Keller, S.; Nentwig, W. Quantity and quality of venom released by a spider (Cupiennius salei, Ctenidae). Toxicon 1995, 33, 1347–1357. [Google Scholar] [CrossRef]
- Malli, H.; Imboden, H.; Kuhn-Nentwig, L. Quantifying the venom dose of the spider Cupiennius salei using monoclonal antibodies. Toxicon 1998, 36, 1959–1969. [Google Scholar] [CrossRef]
- Malli, H.; Kuhn-Nentwig, L.; Imboden, H.; Nentwig, W. Effects of size, motility and paralysation time of prey on the quantity of venom injected by the hunting spider Cupiennius salei. J. Exp. Biol. 1999, 202, 2083–2089. [Google Scholar] [CrossRef]
- Schendel, V.; Rash, L.D.; Jenner, R.A.; Undheim, E.A.B. The Diversity of Venom: The Importance of Behavior and Venom System Morphology in Understanding Its Ecology and Evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef]
- Herzig, V.; King, G.F.; Undheim, E.A. Can we resolve the taxonomic bias in spider venom research? Toxicon X 2019, 1, 100005. [Google Scholar] [CrossRef]
- Vassilevski, A.A.; Kozlov, S.A.; Grishin, E.V. Molecular diversity of spider venom. Biochemistry 2009, 74, 1505–1534. [Google Scholar] [CrossRef]
- Morgenstern, D.; King, G.F. The venom optimization hypothesis revisited. Toxicon 2013, 63, 120–128. [Google Scholar] [CrossRef]
- Dunbar, J.P.; Afoullouss, S.; Sulpice, R.; Dugon, M.M. Envenomation by the noble false widow spider Steatoda nobilis (Thorell, 1875)–Five new cases of steatodism from Ireland and Great Britain. Clin. Toxicol. 2018, 56, 433–435. [Google Scholar] [CrossRef]
- Atakuziev, B.U.; Wright, C.E.; Graudins, A.; Nicholson, G.M.; Winkel, K.D. Efficacy of Australian red-back spider (Latrodectus hasselti) antivenom in the treatment of clinical envenomation by the cupboard spider Steatoda capensis (Theridiidae). Toxicon 2014, 86, 68–78. [Google Scholar] [CrossRef]
- Nentwig, W.; Friedel, T.; Manhart, C. Comparative investigations on the effect of the venoms of 18 spider species onto the cockroach Blatta orientalis (Blattodea). Zool. Jb. Physiol. 1992, 96, 279–290. [Google Scholar]
- Betz, O.; Kölsch, G. The role of adhesion in prey capture and predator defence in arthropods. Arthropod Struct. Dev. 2004, 33, 3–30. [Google Scholar] [CrossRef]
- Blackledge, T.; Coddington, J.; Gillespie, R. Are three-dimensional spider webs defensive adaptations? Ecol. Lett. 2003, 6, 13–18. [Google Scholar] [CrossRef]
- Forster, L. The behavioural ecology of Latrodectus hasselti (Thorell), the Australian redback spider (Araneae: Theridiidae): A review. Rec. West. Aust. Mus. 1995, 52, 13–24. [Google Scholar]
- Anotaux, M.; Toscani, C.; Leborgne, R.; Châline, N.; Pasquet, A. Aging and foraging efficiency in an orb-web spider. J. Ethol. 2014, 32, 155–163. [Google Scholar] [CrossRef]
- Jackson, R.R.; Brassington, R.J. The biology of Pholcus phalangioides (Araneae, Pholcidae): Predatory versatility, araneophagy and aggressive mimicry. J. Zool. 1987, 211, 227–238. [Google Scholar] [CrossRef]
- Roberts, M.J. Collins Field Guide: Spiders of Britain & Northern Europe; Harper Collins: London, UK, 1995. [Google Scholar]
- Herzig, V.; Ward, R.J.; dos Santos, W.F. Intersexual variations in the venom of the Brazilian ′armed′ spider Phoneutria nigriventer (Keyserling, 1891). Toxicon 2002, 40, 1399–1406. [Google Scholar] [CrossRef]
- Vapenik, Z.; Nentwig, W. The influence of hunger and breeding temperature on the venom production of the spider Cupiennius salei (Araneae, Ctenidae). Toxicon 2000, 38, 293–298. [Google Scholar] [CrossRef]
- Binford, G.J. An analysis of geographic and intersexual chemical variation in venoms of the spider Tegenaria agrestis (Agelenidae). Toxicon 2001, 39, 955–968. [Google Scholar] [CrossRef]
- Zobel-Thropp, P.A.; Mullins, J.; Kristensen, C.; Kronmiller, B.A.; David, C.L.; Breci, L.A.; Binford, G.J. Not so dangerous after all? Venom composition and potency of the pholcid (daddy long-leg) spider Physocyclus mexicanus. Front. Ecol. Evol. 2019, 7, 256. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. A practical information-theoretic approach. In Model Selection and Multimodel Inference; Springer: New York, NY, USA, 2002; Volume 2, pp. 70–71. [Google Scholar]
- Venables, B.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sex | Body Size (mm) | Venom Yield (µL) |
|---|---|---|---|
| S. nobilis | Female | 5.70–13.90 | 0.01–0.63 |
| n = 425 | 9.85 ± 1.41 | 0.23 ± 0.11 | |
| Male | 5.40–12.50 | 0.03–0.50 | |
| n = 125 | 9.15 ±1.23 | 0.19 ± 0.10 | |
| A. similis | Female | 6.20–13.00 | 0.01–0.69 |
| n = 187 | 10.02 ± 1.5 | 0.24 ± 0.16 | |
| Male | 5.80–12.00 | 0.01–0.35 | |
| n = 78 | 8.69 ± 1.07 | 0.12 ± 0.08 | |
| E. atrica | Female | 6.00–18.00 | 0.03–1.71 |
| n = 71 | 11.98 ± 2.72 | 0.52 ± 0.40 | |
| Male | 7.10–17.40 | 0.10–2.25 | |
| n = 68 | 12.63 ± 2.50 | 0.75 ± 0.48 |
| Fixed Factors | Estimate | Standard Error | p-Value |
|---|---|---|---|
| Intercept (Baseline: S nobilis, Autumn, Female) | −0.022 | 0.049 | 0.65 |
| Body size (mm) (S. nobilis) | 0.027 | 0.005 | <0.001 |
| Season Spring | −0.029 | 0.019 | 0.18 |
| Season Summer | −0.045 | 0.012 | <0.005 |
| Sex Male | −0.006 | 0.012 | 0.64 |
| Species Intercept (E atrica) | −0.809 | 0.082 | <0.001 |
| Species Intercept (A similis) | −0.132 | 0.081 | 0.10 |
| Interaction Size: E. atrica | 0.095 | 0.007 | <0.001 |
| Interaction Size: A. similis | 0.012 | 0.008 | 0.145 |
| Estimate | Standard Error | p-Value | |
|---|---|---|---|
| Intercept (C2) | 2.198 | 0.745 | <0.05 |
| C1 | 16.369 | - | - |
| L8 | −2.197 | 0.869 | <0.05 |
| L48 | −1.350 | 0.891 | 0.130 |
| S8 | −2.00 | 0.87 | <0.05 |
| S48 | −2.00 | 0.87 | <0.05 |
| A. similis Venom | E. atrica Venom | S. nobilis Venom | ||||
|---|---|---|---|---|---|---|
| ED50, LD50 (mg/kg) | SE | ED50, LD50 (mg/kg) | SE | ED50, LD50 (mg/kg) | SE | |
| Prey | ||||||
| A. similis | N/A | - | ED50 = 10.07 LD50 = 445.30 | 0.26 0.50 | ED50 = 0.25 LD50 = 0.96 | 0.14 90.20 |
| E. atrica | ED50 = 34.40 LD50 > 60.44 | 0.17 - | N/A | - | ED50 = 0.71 LD50 = 0.46 | 0.17 0.23 |
| S. nobilis | ED50 > 98.73 LD50 > 98.73 | - - | ED50 = 4.14 LD50 > 188.17 | 0.53 - | N/A | - |
| Z. x-notata | ED50 = 0.59 LD50 = 128.50 | 1.46 0.27 | ED50 = 2.39 LD50 = 69.80 | 0.31 0.16 | ED50 = 0.09 LD50 = 0.56 | 0.22 0.14 |
| Prey | Venom Concentrations (%) | ||
|---|---|---|---|
| A. similis Venom | E. atrica Venom | S. nobilis Venom | |
| A. similis | N/A | 0.01, 0.1, 1, 2 | 0.0001, 0.001, 0.01, 0.1, 1 |
| E. atrica | 0.01, 0.1, 1, 2 | N/A | 0.01, 0.1, 1 |
| S. nobilis | 0.01, 0.1, 1, 2 | 0.01, 0.1, 1, 2 | N/A |
| Z. x-notata | 0.01, 0.1, 1, 2 | 0.001, 0.01, 0.1, 1 | 0.0001, 0.001, 0.01, 0.1, 1 |
| Cohort | Anaesthetised | Venom Extracted | Time (h) | Prey Size |
|---|---|---|---|---|
| C1 | No | No | N/A | Large |
| C2 | Yes | Electrostimulation only | 8 | Large |
| L8 | Yes | Yes | 8 | Large |
| L48 | Yes | Yes | 48 | Large |
| S8 | Yes | Yes | 8 | Small |
| S48 | Yes | Yes | 48 | Small |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rayner, S.; Vitkauskaite, A.; Healy, K.; Lyons, K.; McSharry, L.; Leonard, D.; Dunbar, J.P.; Dugon, M.M. Worldwide Web: High Venom Potency and Ability to Optimize Venom Usage Make the Globally Invasive Noble False Widow Spider Steatoda nobilis (Thorell, 1875) (Theridiidae) Highly Competitive against Native European Spiders Sharing the Same Habitats. Toxins 2022, 14, 587. https://doi.org/10.3390/toxins14090587
Rayner S, Vitkauskaite A, Healy K, Lyons K, McSharry L, Leonard D, Dunbar JP, Dugon MM. Worldwide Web: High Venom Potency and Ability to Optimize Venom Usage Make the Globally Invasive Noble False Widow Spider Steatoda nobilis (Thorell, 1875) (Theridiidae) Highly Competitive against Native European Spiders Sharing the Same Habitats. Toxins. 2022; 14(9):587. https://doi.org/10.3390/toxins14090587
Chicago/Turabian StyleRayner, Sean, Aiste Vitkauskaite, Kevin Healy, Keith Lyons, Leona McSharry, Dayle Leonard, John P. Dunbar, and Michel M. Dugon. 2022. "Worldwide Web: High Venom Potency and Ability to Optimize Venom Usage Make the Globally Invasive Noble False Widow Spider Steatoda nobilis (Thorell, 1875) (Theridiidae) Highly Competitive against Native European Spiders Sharing the Same Habitats" Toxins 14, no. 9: 587. https://doi.org/10.3390/toxins14090587